Gnaiger 2015 Scand J Med Sci Sports

| Gnaiger E, Boushel R, Søndergaard H, Munch-Andersen T, Damsgaard R, Hagen C, Díez-Sánchez C, Ara I, Wright-Paradis C, Schrauwen P, Hesselink M, Calbet JAL, Christiansen M, Helge JW, Saltin B (2015) Mitochondrial coupling and capacity of oxidative phosphorylation in skeletal muscle of Inuit and caucasians in the arctic winter. https://doi.org/10.1111/sms.12612 |
» Scand J Med Sci Sports 25 (Suppl 4):126–34. PMID: 26589126 Open Access »![]()
Gnaiger Erich, Boushel R, Soendergaard H, Munch-Andersen T, Damsgaard R, Hagen C, Diez-Sanchez C, Ara I, Wright-Paradis C, Schrauwen P, Hesselink M, Calbet Jose AL, Christiansen M, Helge JW, Saltin Bengt (2015) Scand J Med Sci Sports
Abstract:  O2k-in brief
O2k-in brief
During evolution, mtDNA haplogroups of arctic populations may have been selected for lower coupling of mitochondrial respiration to ATP production in favor of higher heat production. We show that mitochondrial coupling in skeletal muscle of traditional and westernized Inuit habituating northern Greenland is identical to Danes of western Europe haplogroups. Biochemical coupling efficiency was preserved across variations in diet, muscle fibre type and uncoupling protein-3 content. Mitochondrial phenotype displayed plasticity in relation to lifestyle and environment. Untrained Inuit and Danes had identical capacities to oxidize fat substrate in arm muscle, which increased in Danes during the 42 days of acclimation to exercise, approaching the higher level of the Inuit hunters. A common pattern emerges of mitochondrial acclimatization and evolutionary adaptation in humans at high latitude and high altitude where economy of locomotion may be optimized by preservation of biochemical coupling efficiency at modest mitochondrial density, when submaximum performance is uncoupled from VO2max and maximum capacities of oxidative phosphorylation.
• Keywords: BMI, VO2max
• Bioblast editor: Gnaiger E
• O2k-Network Lab: AT Innsbruck Gnaiger E, AT Innsbruck Oroboros, CA Vancouver Boushel RC, NL Maastricht Schrauwen P, DK Copenhagen Christiansen M, ES CN Las Palmas Calbet JAL
SUIT protocol


Greenland expedition CMRC: science and adventure
MitoEAGLE VO2max/BME database
- Human vastus lateralis
- 8 females & 8 males
- 31 years
- Active; Danes, baseline control in Copenhagen
- H = 1.75 m
- M = 74.5 kg
- BME = 0.19
- BMI = 24.3 kg·m-2
- VO2max/M = 48.0 mL·min-1·kg-1
- Permeabilized muscle fibres; 30 °C; GMSP; mw; conversions: Gnaiger 2009 Int J Biochem Cell Biol
- JO2,P(NS) = 90.5 µmol·s-1·kg-1 wet muscle mass (37 °C)
Bengt Saltin - a Gentle Scientist

Additional references
- Marconi C, Marzorati M, Cerretelli P (2006) Work capacity of permanent residents of high altitude. High Alt Med Biol 7:105-115. - »Bioblast link«
- Tam E, Bruseghini P, Calabria E, Sacco LD, Doria C, Grassi B, Pietrangelo T, Pogliaghi S, Reggiani C, Salvadego D, Schena F, Toniolo L, Verratti V, Vernillo G, Capelli C (2015) Gokyo Khumbu/Ama Dablam Trek 2012: effects of physical training and high-altitude exposure on oxidative metabolism, muscle composition, and metabolic cost of walking in women. Eur J Appl Physiol. - »Bioblast link«
O2k-brief
MitoFit news 2015#18
- 2015-11-25: Performance tests on skeletal muscle mitochondria of the Inuit haplogroup reveal fitness information beyond the uncoupling hypothesis for adaptations to the arctic climate. » MitoFit news - a contribution to K-Regio MitoFit.
References: BME and VO2max
- » VO2max
| Reference | |
|---|---|
| Bakkman 2007 ActaPhysiol | Bakkman L, Sahlin K, Holmberg HC, Tonkonogi M (2007) Quantitative and qualitative adaptation of human skeletal muscle mitochondria to hypoxic compared with normoxic training at the same relative work rate. Acta Physiol (Oxford) 190:243–51. |
| Boushel 2007 Diabetologia | Boushel RC, Gnaiger E, Schjerling P, Skovbro M, Kraunsoee R, Dela F (2007) Patients with Type 2 diabetes have normal mitochondrial function in skeletal muscle. Diabetologia 50:790-6. |
| Chambers 2020 J Appl Physiol (1985) | Chambers TL, Burnett TR, Raue U, Lee GA, Finch WH, Graham BM, Trappe TA, Trappe S (2020) Skeletal muscle size, function, and adiposity with lifelong aerobic exercise. J Appl Physiol (1985) 128:368–78. |
| Daussin 2008 Am J Physiol Regul Integr Comp Physiol | Daussin FN, Zoll J, Dufour SP, Ponsot E, Lonsdorfer-Wolf E, Doutreleau S, Mettauer B, Piquard F, Geny B, Richard R (2008) Effect of interval versus continuous training on cardiorespiratory and mitochondrial functions: relationship to aerobic performance improvements in sedentary subjects. Am J Physiol Regul Integr Comp Physiol 295:R264-72. |
| Garnier 2005 FASEB J | Garnier A, Fortin D, Zoll J, N'Guessan B, Mettauer B, Lampert E, Veksler V, Ventura-Clapier R (2005) Coordinated changes in mitochondrial function and biogenesis in healthy and diseased human skeletal muscle. FASEB J 19:43-52. |
| Gnaiger 2015 Scand J Med Sci Sports | Gnaiger E, Boushel R, Søndergaard H, Munch-Andersen T, Damsgaard R, Hagen C, Díez-Sánchez C, Ara I, Wright-Paradis C, Schrauwen P, Hesselink M, Calbet JAL, Christiansen M, Helge JW, Saltin B (2015) Mitochondrial coupling and capacity of oxidative phosphorylation in skeletal muscle of Inuit and caucasians in the arctic winter. https://doi.org/10.1111/sms.12612 |
| Gnaiger 2019 MiP2019 |  |
| Loe 2013 PLOS ONE | Loe H, Rognmo Ø, Saltin B, Wisløff U (2013) Aerobic capacity reference data in 3816 healthy men and women 20-90 years. PLOS ONE 8:e64319. |
| Mettauer 2001 J Am Coll Cardiol | Mettauer B, Zoll J, Sanchez H, Lampert E, Ribera F, Veksler V, Bigard X, Mateo P, Epailly E, Lonsdorfer J, Ventura-Clapier R (2001) Oxidative capacity of skeletal muscle in heart failure patients versus sedentary or active control subjects. J Am Coll Cardiol 38:947-54. |
| Mogensen 2006 J Physiol | Mogensen M, Bagger M, Pedersen PK, Fernström M, Sahlin K (2006) Cycling efficiency in humans is related to low UCP3 content and to type I fibres but not to mitochondrial efficiency. J Physiol 571:669-81. |
| N'Guessan 2004 Mol Cell Biochem | N'Guessan B, Zoll J, Ribera F, Ponsot E, Lampert E, Ventura-Clapier R, Veksler V, Mettauer B (2004) Evaluation of quantitative and qualitative aspects of mitochondrial function in human skeletal and cardiac muscles. Mol Cell Biochem 256-257:267-80. |
| Pesta 2011 Am J Physiol Regul Integr Comp Physiol | Pesta D, Hoppel F, Macek C, Messner H, Faulhaber M, Kobel C, Parson W, Burtscher M, Schocke M, Gnaiger E (2011) Similar qualitative and quantitative changes of mitochondrial respiration following strength and endurance training in normoxia and hypoxia in sedentary humans. Am J Physiol Regul Integr Comp Physiol 301:R1078–87. |
| Ponsot 2006 J Appl Physiol (1985) | Ponsot E, Dufour SP, Zoll J, Doutrelau S, N'Guessan B, Geny B, Hoppeler H, Lampert E, Mettauer B, Ventura-Clapier R, Richard R (2006) Exercise training in normobaric hypoxia in endurance runners. II. Improvement of mitochondrial properties in skeletal muscle. J Appl Physiol (1985) 100:1249-57. |
| Pribis 2010 Nutrients | Pribis P, Burtnack CA, McKenzie SO, Thayer J (2010) Trends in body fat, body mass index and physical fitness among male and female college students. Nutrients 2:1075-85. |
| Raboel 2009 Diabetes Obes Metab | Raboel R, Hojberg PM, Almdal T, Boushel RC, Haugaard SB, Madsbad S, Dela F (2009) Improved glycaemic control decreases inner mitochondrial membrane leak in type 2 diabetes. Diabetes Obes Metab 11:355-60. |
| Rasmussen 2001 Am J Physiol Endocrinol Metab | Rasmussen UF, Rasmussen HN, Krustrup P, Quistorff B, Saltin B, Bangsbo J (2001) Aerobic metabolism of human quadriceps muscle: in vivo data parallel measurements on isolated mitochondria. Am J Physiol Endocrinol Metab 280:E301-7. |
| Rasmussen 2003 Eur J Physiol | Rasmussen UF, Krustrup P, Kjaer M, Rasmussen HN (2003) Human skeletal muscle mitochondrial metabolism in youth and senescence: no signs of functional changes in ATP formation and mitochondrial oxidative capacity. Pflugers Arch – Eur J Physiol 446:270-78. |
| Zoll 2002 J Physiol | Zoll J, Sanchez H, N'Guessan B, Ribera F, Lampert E, Bigard X, Surrurier B, Fortin D, Geny B, Veksler V, Ventura-Clapier R, Mettauer B (2002) Physical activity changes the regulation of mitochondrial respiration in human skeletal muscle. J Physiol 543:191-200. |
MitoPedia: BME and mitObesity
» Body mass excess and mitObesity | BME and mitObesity news | Summary |
| Term | Abbreviation | Description |
|---|---|---|
| BME cutoff points | BME cutoff | Obesity is defined as a disease associated with an excess of body fat with respect to a healthy reference condition. Cutoff points for body mass excess, BME cutoff points, define the critical values for underweight (-0.1 and -0.2), overweight (0.2), and various degrees of obesity (0.4, 0.6, 0.8, and above). BME cutoffs are calibrated by crossover-points of BME with established BMI cutoffs. |
| Body fat excess | BFE | In the healthy reference population (HRP), there is zero body fat excess, BFE, and the fraction of excess body fat in the HRP is expressed - by definition - relative to the reference body mass, M°, at any given height. Importantly, body fat excess, BFE, and body mass excess, BME, are linearly related, which is not the case for the body mass index, BMI. |
| Body mass | m [kg]; M [kg·x-1] | The body mass M is the mass (kilogram [kg]) of an individual (object) [x] and is expressed in units [kg/x]. Whereas the body weight changes as a function of gravitational force (you are weightless at zero gravity; your floating weight in water is different from your weight in air), your mass is independent of gravitational force, and it is the same in air and water. |
| Body mass excess | BME | The body mass excess, BME, is an index of obesity and as such BME is a lifestyle metric. The BME is a measure of the extent to which your actual body mass, M [kg/x], deviates from M° [kg/x], which is the reference body mass [kg] per individual [x] without excess body fat in the healthy reference population, HRP. A balanced BME is BME° = 0.0 with a band width of -0.1 towards underweight and +0.2 towards overweight. The BME is linearly related to the body fat excess. |
| Body mass index | BMI | The body mass index, BMI, is the ratio of body mass to height squared (BMI=M·H-2), recommended by the WHO as a general indicator of underweight (BMI<18.5 kg·m-2), overweight (BMI>25 kg·m-2) and obesity (BMI>30 kg·m-2). Keys et al (1972; see 2014) emphasized that 'the prime criterion must be the relative independence of the index from height'. It is exactly the dependence of the BMI on height - from children to adults, women to men, Caucasians to Asians -, which requires adjustments of BMI-cutoff points. This deficiency is resolved by the body mass excess relative to the healthy reference population. |
| Comorbidity | Comorbidities are common in obesogenic lifestyle-induced early aging. These are preventable, non-communicable diseases with strong associations to obesity. In many studies, cause and effect in the sequence of onset of comorbidities remain elusive. Chronic degenerative diseases are commonly obesity-induced. The search for the link between obesity and the etiology of diverse preventable diseases lead to the hypothesis, that mitochondrial dysfunction is the common mechanism, summarized in the term 'mitObesity'. | |
| Healthy reference population | HRP | A healthy reference population, HRP, establishes the baseline for the relation between body mass and height in healthy people of zero underweight or overweight, providing a reference for evaluation of deviations towards underweight or overweight and obesity. The WHO Child Growth Standards (WHO-CGS) on height and body mass refer to healthy girls and boys from Brazil, Ghana, India, Norway, Oman and the USA. The Committee on Biological Handbooks compiled data on height and body mass of healthy males from infancy to old age (USA), published before emergence of the fast-food and soft-drink epidemic. Four allometric phases are distinguished with distinct allometric exponents. At heights above 1.26 m/x the allometric exponent is 2.9, equal in women and men, and significantly different from the exponent of 2.0 implicated in the body mass index, BMI [kg/m2]. |
| Height of humans | h [m]; H [m·x-1] | The height of humans, h, is given in SI units in meters [m]. Humans are countable objects, and the symbol and unit of the number of objects is N [x]. The average height of N objects is, H = h/N [m/x], where h is the heights of all N objects measured on top of each other. Therefore, the height per human has the unit [m·x-1] (compare body mass [kg·x-1]). Without further identifyer, H is considered as the standing height of a human, measured without shoes, hair ornaments and heavy outer garments. |
| Length | l [m] | Length l is an SI base quantity with SI base unit meter m. Quantities derived from length are area A [m2] and volume V [m3]. Length is an extensive quantity, increasing additively with the number of objects. The term 'height' h is used for length in cases of vertical position (see height of humans). Length of height per object, LUX [m·x-1] is length per unit-entity UX, in contrast to lentgth of a system, which may contain one or many entities, such as the length of a pipeline assembled from a number NX of individual pipes. Length is a quantity linked to direct sensory, practical experience, as reflected in terms related to length: long/short (height: tall/small). Terms such as 'long/short distance' are then used by analogy in the context of the more abstract quantity time (long/short duration). |
| MitObesity drugs | Bioactive mitObesity compounds are drugs and nutraceuticals with more or less reproducible beneficial effects in the treatment of diverse preventable degenerative diseases implicated in comorbidities linked to obesity, characterized by common mechanisms of action targeting mitochondria. | |
| Obesity | Obesity is a disease resulting from excessive accumulation of body fat. In common obesity (non-syndromic obesity) excessive body fat is due to an obesogenic lifestyle with lack of physical exercise ('couch') and caloric surplus of food consumption ('potato'), causing several comorbidities which are characterized as preventable non-communicable diseases. Persistent body fat excess associated with deficits of physical activity induces a weight-lifting effect on increasing muscle mass with decreasing mitochondrial capacity. Body fat excess, therefore, correlates with body mass excess up to a critical stage of obesogenic lifestyle-induced sarcopenia, when loss of muscle mass results in further deterioration of physical performance particularly at older age. | |
| VO2max | VO2max; VO2max/M | Maximum oxygen consumption, VO2max, is and index of cardiorespiratory fitness, measured by spiroergometry on human and animal organisms capable of controlled physical exercise performance on a treadmill or cycle ergometer. VO2max is the maximum respiration of an organism, expressed as the volume of O2 at STPD consumed per unit of time per individual object [mL.min-1.x-1]. If normalized per body mass of the individual object, M [kg.x-1], mass specific maximum oxygen consumption, VO2max/M, is expressed in units [mL.min-1.kg-1]. |
Cited by
- 13 articles in PubMed (2021-12-27) https://pubmed.ncbi.nlm.nih.gov/26589126/

- Gnaiger E (2020) Mitochondrial pathways and respiratory control. An introduction to OXPHOS analysis. 5th ed. Bioenerg Commun 2020.2. https://doi.org/10.26124/bec:2020-0002
Labels: MiParea: Respiration, mt-Biogenesis;mt-density, mtDNA;mt-genetics, Comparative MiP;environmental MiP, Gender, Exercise physiology;nutrition;life style
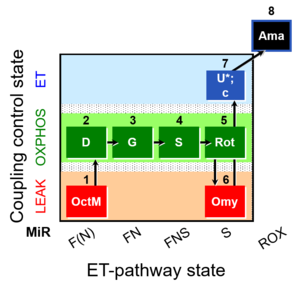
Stress:Temperature Organism: Human Tissue;cell: Skeletal muscle Preparation: Permeabilized tissue Enzyme: Marker enzyme, TCA cycle and matrix dehydrogenases Regulation: Coupling efficiency;uncoupling, Cyt c Coupling state: LEAK, OXPHOS, ET Pathway: F, N, S, ROX HRR: Oxygraph-2k, O2k-Protocol
MitoFitPublication, VO2max, BMI, BME, 1OctM;2D;3G;4S;5Rot;6Omy;7U-, SUIT-016, SUIT-016 O2 pfi D044, MitoEAGLE BME, O2k-brief, BEC 2020.2